

Introduction
The 2017 National Beef Quality Audit reported that the average carcass weight has been increasing at a rate of 1.8 kg per year since 1991 (Boykin et al., 2017). The average carcass weight in 1991 was 345.0 kg, whereas in 2016 it was 390.3 kg (Boykin et al., 2017). This increase in carcass weight poses several challenges for beef processors. For example, the carcass coolers must dissipate considerably higher amounts of heat from larger-framed and heavier-weighing carcasses (McKenna et al., 2002; Boykin et al., 2017). Moreover, with considerably greater amounts of product requiring cooling, quality issues associated with color, tenderness, and water-holding capacity could occur (Savell et al., 2005). Along with these, issues such as sour knuckles are starting to appear with greater frequency in processing plants.
The sour knuckle condition in beef, also referred to as “sour rounds” in some of the literature, is associated with vastus femoris, vastus lateralis, vastus medialis, and rectus femoris muscles expressing a sour/pungent odor that varies in intensity (Haines, 1941; Shank et al., 1962; James and James, 2002; Corry, 2007). It is considered similar to bone souring in hams due to similar anatomical location, and similar Clostridium spp. that have been isolated from both these defects (Ingram, 1952; Cosnett et al., 1956; Nottingham, 1960). On the other hand, some studies have noted that unlike bone souring in hams, the femur marrow is never soured in sour beef knuckles, with the defect being solely located in the muscle tissues alongside the femur (Lepovetsky et al., 1953). In the beef production line setting, this sour condition, reported to be sporadic and unpredictable, is immediately identified when fabricating postchilled beef rounds upon separating the patella from the femur (Gill, 1979; De Lacy et al., 1998).
The majority of studies that have investigated the sour beef round condition were conducted several decades ago, with most indicating a microbial cause for the condition (Haines and Scott, 1940; Lepovetsky et al., 1953; Cosnett et al., 1956; Nottingham, 1960; Shank et al., 1962). However, some studies have reported contradictory findings regarding the contributions of bacterial contamination in soured rounds: some found tissues almost completely sterile, while others found bacterial contamination in sampled meat tissue and synovial fluid (Haines and Scott, 1940; Lepovetsky et al., 1953; Cosnett et al., 1956; Nottingham, 1960; Shank et al., 1962; Gill, 1979). Based on varying intensity of the sourness—which has been described as sweet, sour, and even an “evil-smelling” odor (Nottingham, 1960; Shank et al., 1962)—Nottingham (1960) hypothesized that a mixture of several microorganisms could be responsible for the condition rather than a single microorganism. In a study conducted almost 40 y later, it was demonstrated that the souring condition could be induced using psychrotrophic Clostridium spp. strains previously isolated from spoiled vacuum-packaged meat products (De Lacy et al., 1998).
It has also been hypothesized that the sour beef knuckle condition is not due to microbial contamination, but rather that it originates from complications in the postmortem metabolism (Shank et al., 1962). In support, volatile acid analysis indicated higher amounts of propionic, acetic, and butyric acids in sour rounds compared to non-sour rounds (Shank et al., 1962). Furthermore, Shank et al. (1962) were able to induce the souring condition in lamb legs with improper cooling (37°C overnight, followed by 4°C for 5 d). In addition, a gradient of souring intensities could be recreated by either incorporating propionate into the bloodstream or by extraneous exercise of the ewes prior to slaughter (Shank et al., 1962).
Despite the investigations conducted to date, the cause of the sour knuckle condition remains uncertain (James and James, 2002). In addition, as previously indicated, most of the published studies were conducted several years ago. Since then, many changes have occurred in the beef processing system, and the sour knuckle condition is starting to appear with greater frequency in beef processing plants. Therefore, the objective of the current study was to evaluate the sensory aspects, volatile acids, and microbial populations associated with the sour knuckle condition in beef carcasses to further investigate the etiology of the condition.
Materials and Methods
Sample collection
Beef knuckles (N = 30; vastus femoris, vastus lateralis, vastus medialis, and rectus femoris muscles) were collected during fabrication, over a 2-d period, from a commercial fed-beef processing facility. During the fabrication process, the souring condition was identified by the employees working in the fabrication line. Upon identification, the knuckles were separated into 2 categories based on the intensity of the souring condition, namely, slight sour odor (SLI-SO; knuckles that had a lesser degree of souring expressing dairy sour odor notes) and severe sour odor (SVR-SO; knuckles that had a greater degree of souring expressing raw sewage–like odor notes). Additionally, knuckles that did not express any off-odors were collected randomly during the sample collection and were designated as control samples. A total of 10 knuckles per treatment group were collected over the 2 collection days. Coinciding with collection of each knuckle, sterile polyurethane sponges hydrated with 10 mL of HiCap neutralizing broth (Whirl-Pak; Nasco, Fort Atkinson, WI) were used to separately sample the synovial fluid from where the femur meets the patella, as well as the femur surface.
At the end of each of the 2 collection days, 15 knuckles (5 knuckles per treatment) and corresponding sponges of the synovial fluid and femur surface were transported on ice to the Center for Meat Safety & Quality at Colorado State University (Fort Collins, CO). Upon arrival, knuckles were aseptically halved with one portion containing the vastus lateralis, vastus intermedius, and rectus femoris, while the other contained the vastus intermedius, rectus femoris, and vastus medialis muscles. Halves were randomly assigned to be evaluated within 12 h of collection (day 0) or after 35 d of storage at 0°C ± 2°C under vacuum-packaged conditions (day 35; standard barrier nylon polyethylene bags with 0.6 cm3 O2/645.16 cm2/24 h at 0°C). Sponge samples were shipped overnight to a commercial testing laboratory for microbiological analysis.
Odor panels
Odor panelists (n = 7) were trained prior to day 0 and day 35 of odor panels. Panel anchors for oxidation and sour notes were trained with references set by Adhikari et al. (2010). In addition, putrid notes and overall off-odor were described and given anchors to train the panelists (Table 1). At the start of each day’s odor panel, anchors were reintroduced before giving the panelists 5 knuckle samples per treatment group (control, SLI-SO, SVR-SO) to evaluate. On each of the odor panel days (day 0 and day 35), 15 samples of muscle tissue originally in contact with the femur (5 of each treatment group) were diced into 1.27 cm ×2.5 cm, and 3 pieces were placed into glass test tubes (55 mL; 25 mm × 150 mm) labeled with randomized 3-digit codes. The test tubes were immediately capped and held at 0°C ± 2°C (1–3 h) until the start of the odor panel. Approximately 20 min before the panels, samples were taken out of the refrigerator and held at room temperature (22°C ± 2°C) to allow volatile compounds to enter the headspace of the test tubes. During testing, panelists were housed in individual sensory booths and were provided with a vortex, test tube rack, and a nasal palate cleanser (coffee beans) to use between samples. Panelists were instructed to vortex the samples in the test tubes for 5 s prior to odor evaluation, and to utilize the nasal palate cleanser between samples with 1 min of rest between each sample. The results were reported on a 10-point line scale through an online survey software (Qualtrics, Provo, UT). In the ballot, panelists were given a line scale to rate all 4 of the attributes (sour, putrid, oxidative, and overall off-odors)—with a 0 representing no sour, no putrid, no oxidation, and no overall-off odors; a 5 representing unacceptable sour, unacceptable putrid, unacceptable oxidized, and unacceptable off-odors; and a 10 representing extremely sour, extremely putrid, extremely oxidative, and extreme off-odors.
Definitions and references for beef knuckle odor panel notes evaluated
| Attribute | Definition | Reference |
| Oxidation | The aromatics commonly associated with oxidized fat and oils. These aromatics may include cardboard, painty, varnish, and fishy.* | Wesson vegetable oil = 1.0 Microwaved Wesson vegetable oil (3 min at high) = 8.0 |
| Putridity | The aromatics associated with spoiled meat | Fresh beef NY strip steak = 0.0 Beef NY strip steak held at 22°C ± 2°C for 24 h = 6.0 |
| Sour | Sour, fermented aromatics associated with dairy products such as buttermilk and sour cream* | Fresh sour cream = 2.0 Sour cream held at 22°C ± 2°C for 24 h = 5.0 |
| Overall off-odor | The combination of sour, putrid, and oxidative notes | Non-sour knuckle = 2.0 Putrid knuckle collected from processing facility = 7.0 |
Refer to Adhikari et al. (2010) for further details.
Gas chromatography-mass spectrometry
Gas chromatography-mass spectrometry (GC-MS) analysis was only performed on day 0 of storage (i.e., day of sample collection) and only on control and SVR-SO (severely sour) samples to examine the 2 extremes. A portion (5 g) of the meat tissue originally in contact with the femur was diced into small cubes and frozen with liquid nitrogen before being transferred into a 20 mL headspace vial and stored at −80°C until analysis. For the GC-MS, samples were incubated at 40°C for 30 min followed by extraction using a Carboxen/PDMS solid phase microextraction fiber (85 µm; Stableflex, Sigma-Aldrich, St. Louis, MO), as described by Pérez et al. (2008). Following extraction, compounds were injected into a DB-WAXUI column (30 m × 0.25 μm; Agilent Technologies, Santa Clara, CA) in a TRACE 1310 GC (Thermo Fisher Scientific, Waltham, MA) coupled to an ISQ-LT MS (Thermo Fisher Scientific). The solid phase microextraction fiber was desorbed at the injection port (250°C) for 3 min, and additionally at the fiber conditioning port (270°C) for 3 min with the GC inlet operating under splitless mode. The oven program started at 35°C for 5 min, with the first ramp to 100°C at a rate of 8°C/min, the second ramp to 240°C at a rate of 12°C/min, and a final hold at 240°C for 5 min. These data were collected under the electron impact mode with a full scan of 35–350 atomic mass units at a scan rate of 10 scans/s.
Microbiological analyses
Beef knuckle muscle tissue that was previously in direct contact with the femur was analyzed for aerobic plate counts (APC) and lactic acid bacteria counts (LABC) on day 0 and day 35 of storage. For each sample, 15 g of muscle tissue was aseptically excised and placed into a Whirl-Pak filter bag (24 oz; Nasco) together with 30 mL of maximum recovery diluent (Neogen, Lansing, MI). Samples were mechanically pummeled (Masticator, IUL Instruments, Barcelona, Spain) for 2 min and diluted in 0.1% buffered peptone water (Difco, Becton Dickinson and Company [BD], Sparks, MD). Appropriate dilutions were then plated, in duplicate, onto Petrifilm Aerobic Count Plates (3M, St. Paul, MN) for determination of APC. To obtain LABC for the samples, 1 mL of appropriate dilutions was transferred to empty petri dishes, in duplicate. Then, 10 mL of molten (45°C–50°C) Lactobacilli MRS agar (Difco, BD) was added to each petri dish, swirled, and allowed to set at room temperature. After the agar was set, a 10 mL overlay of molten Lactobacilli MRS agar was added to each plate. Colonies were counted after incubation of plates at 25°C ± 2°C for 72 h (Petrifilm Aerobic Count Plate) or 5 d (Lactobacilli MRS agar). The detection limit was 0.5 log colony forming units (CFU)/g for both analyses (i.e., APC and LABC).
Sponge samples of the synovial fluid and femur surface were analyzed for psychrotrophic anaerobic sporeformer counts. The procedure followed was based on the methodology described for mesophilic anaerobic sporeformers in the Compendium of Methods for the Microbiological Examination of Foods (Scott et al., 2001), with a modification made to the incubation conditions (temperature and duration) of the agar plates so as to detect psychrotrophic anaerobic sporeformers (Broda et al., 1998). Briefly, upon arrival at the commercial testing laboratory, 15 mL of peptone water diluent was added to each sponge, followed by mechanical pummeling (60 s). An aliquot (10 mL) of sample was then removed and added to a sterile flask containing 90 mL of tempered (approximately 45°C ± 2°C) molten tryptone glucose extract agar (Neogen). Flasks were gently agitated and immediately placed in a circulating water bath set at 80°C ± 1°C. They were held at this temperature for 30 min with occasional agitation. Following incubation, flasks were cooled in tepid (45°C ± 2°C) water for ≤10 min before being poured into 5 petri dishes (approximately 20 mL per plate). Upon solidification of the agar, plates were incubated anaerobically (GasPak EZ Anaerobe Container System; BD) at 25°C ± 2°C for 10 d (Broda et al., 1998). After the incubation period, colonies exhibiting typical morphology were counted, and the number of CFU per sponge determined. The detection limit of the analysis was 0.4 log CFU/sponge.
Statistical analysis
Bacterial populations (APC and LABC) recovered from muscle tissue samples were expressed as least-squares means for log CFU/g, and psychrotrophic anaerobic sporeformer counts recovered from sponge samples of the synovial fluid and femur surface were expressed as least-squares means for log CFU/sponge. For the purpose of statistical analysis of the microbial data, samples with counts below the analysis detection limit were assigned a value equal to the detection limit (i.e., 0.5 log CFU/g for APC and LABC, and 0.4 log CFU/sponge for psychrotrophic anaerobic sporeformer counts). Data analysis of odor panels and microbial populations were conducted as a paired comparison design. Odor panel scores for all tested attributes, APC, LABC, and psychrotrophic anaerobic sporeformer counts were analyzed using R Studio (version 3.5.1), with treatment (control, SLI-SO, or SVR-SO) as the factor of the study design with significance at an alpha level of 0.05. The analysis of variance function was utilized from R Studio to determine significant differences. Upon identification of a significant difference (P < 0.05), the emmeans function was used to determine the statistical difference between treatment groups.
For each GC-MS sample, raw data files were converted to. cdf format, and matrix of molecular features as defined by retention time and mass (m/z) was generated using XCMS software in R for feature detection and alignment (Smith et al., 2006). The matchedFilter algorithm was used for GC-MS data. Features were grouped using RAMClustR (Broeckling et al., 2014), with options set as hmax = 0.9 min, ModuleSize = 3, linkage = “average,” and normalize = “TIC.” Compounds were annotated based on spectral matching to NIST version 12. Principal component analysis was conducted on mean-centered and pareto variance-scaled data using the pcaMethods package in R. Analysis of variance was conducted on each compound using the aov function in R, and P values were adjusted for false positives using the Bonferroni-Hochberg method in the p.adjust function in R (Benjamini and Hochberg, 1995).
Results and Discussion
Odor panels
Odor panels performed on knuckles on day 0 indicated differences (P < 0.05) for off-odor, oxidation, putrid, and sour notes between control and sour knuckles (both SLI-SO and SVR-SO; Figure 1A), with sour knuckles having a greater (P < 0.05) intensity for all attributes tested. However, the panelists were not able (P > 0.05) to differentiate between SLI-SO and SVR-SO knuckles on day 0 for all the evaluated attributes (Figure 1A). For samples stored for 35 d, differences (P < 0.05) were observed between control, SLI-SO, and SVR-SO knuckles for off-odor, oxidation, putrid, and sour notes, with SVR-SO having the greatest values (P < 0.05) for all attributes tested (Figure 1B). Under the current meat processing practices in the United States, there are chances for a soured knuckle to enter the marketplace as most of the processing plants are relying on the subjective judgement of an employee in the fabrication line to identify this quality defect. However, it is unlikely to reach the customers on the same day (i.e., day 0, in the current study). Considering the average of 20.5 d (ranging from 1 to 358 d) for a beef product to reach the retail market (Guelker et al., 2013), products exhibiting the souring condition in the retail setting might resemble results from day 35 panels obtained in the current study, with the retailer or consumer being able to differentiate between a slight and severely sour knuckle. In the current study, we evaluated the products on day 35, which is usually the upper limit for refrigerated storage of products in vacuum (Blixt and Borch, 2002). If the sour knuckles enter the retail distribution chain, the producers risk negative perceptions for their products, thereby harming retailer or consumer trust. While the sour odors have been described to range from sweet to sewer-like, further research is needed to determine the thresholds for retailer/consumer rejection for the soured products (Nottingham, 1960; Shank et al., 1962).
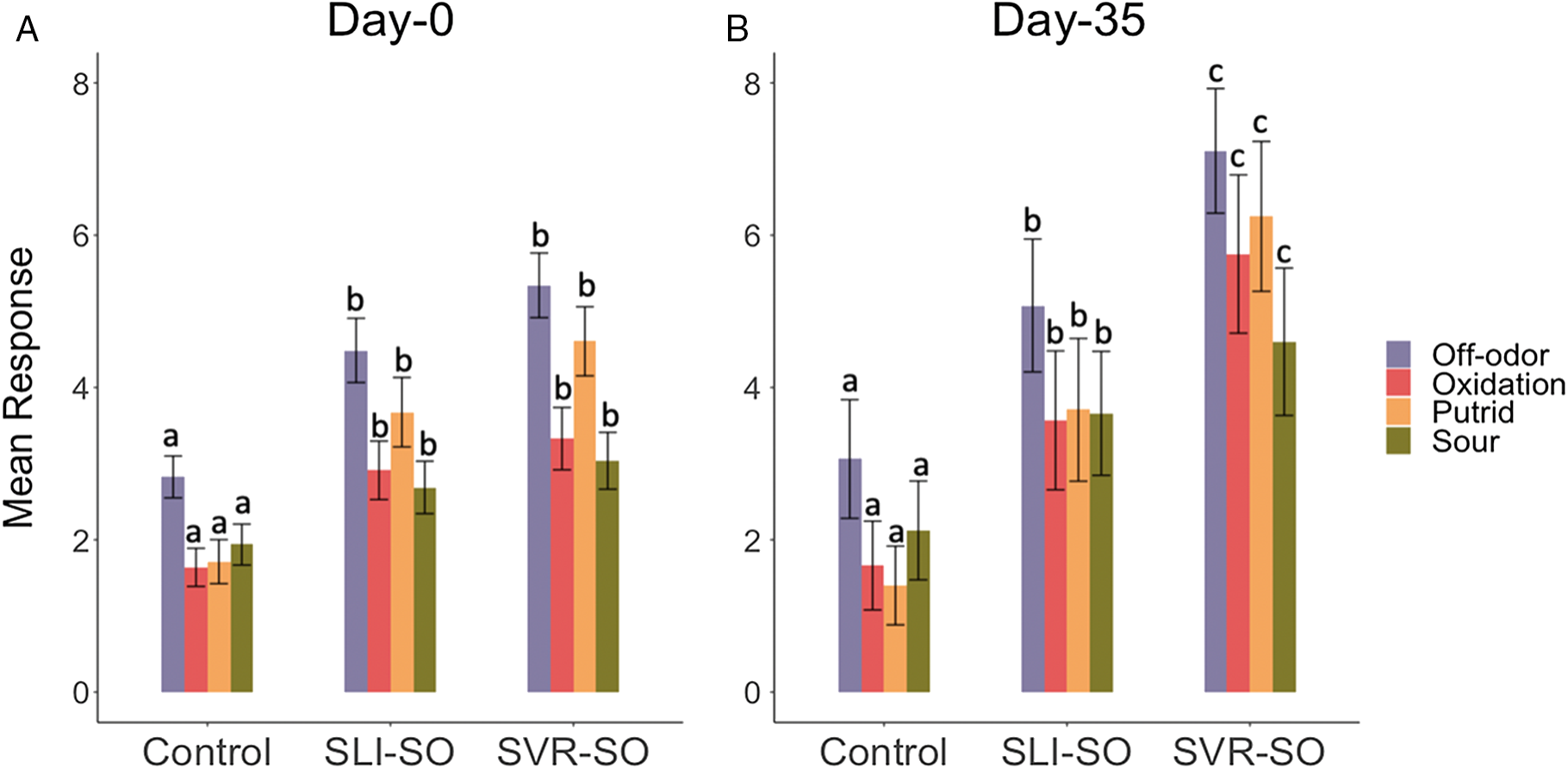
Mean responses from trained odor panelists (n = 7) for off-odor, oxidation, putrid, and sour notes of knuckles (n = 10) with no sour odor (control), slight sour odor (SLI-SO), and severe sour odor (SVR-SO) on (A) day 0 and (B) day 35 following vacuum-packaged storage at 0°C ± 2°C. a–cLetters within each storage day and attribute with different letter assignments differ statistically (P < 0.05).
GC-MS
Although the odor panelists were able to identify the difference in sourness between treatments, GC-MS results indicated no statistical difference (P > 0.05) between control and SVR-SO samples on day 0. These results were unexpected as the GC-MS analysis was only performed on control and the severely sour (SVR-SO) treatment groups to compare the two extremes. Moreover, Shank et al. (1962) reported that sour rounds had greater amounts of formic, acetic, propionic, and butyric acids compared to non-sour controls. In the current study, a wide variation was observed among samples within each of the treatment groups when evaluating acetoin, and acetic, butyric, and propionic acids (Figure 2). These results suggest that the extent of sourness was not consistent among samples within the treatment group and could have possibly led to the lack of statistical differences.
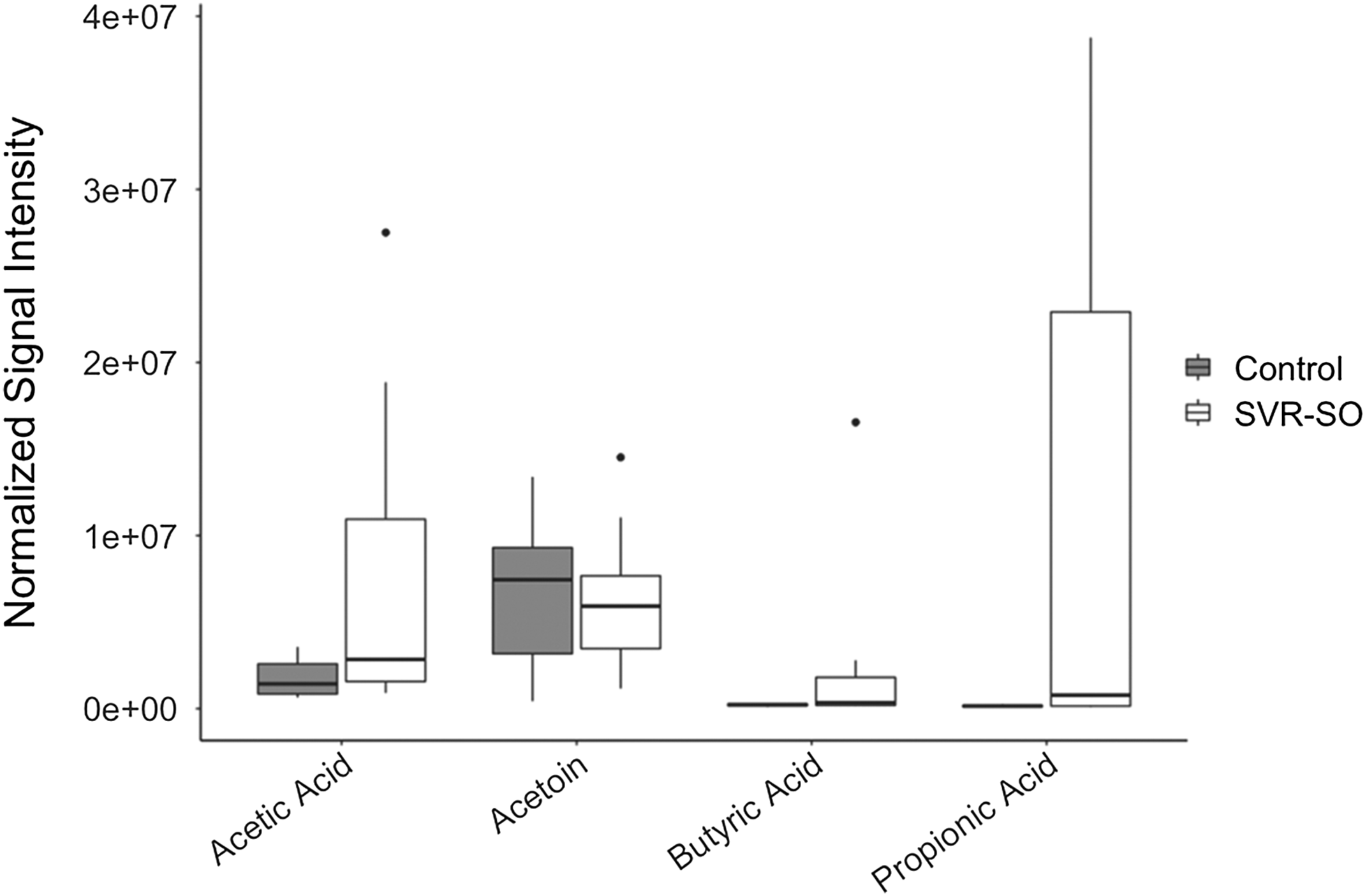
Normalized abundance averages (n = 10) for the major short chain fatty acids identified from beef knuckles with no sour odor (control) and severe sour odor (SVR-SO) on day 0 using gas chromatography-mass spectrometry. The dots/circles represent outliers.
Microbial levels of knuckle muscle tissue
Muscle tissue from control, SLI-SO, and SVR-SO knuckles was analyzed for APC and LABC to determine whether there were differences between sour and non-sour knuckles for these microbiological indicators of meat quality. Results showed similar (P > 0.05) APC and LABC for control, SLI-SO, and SVR-SO samples, irrespective of sampling day (day 0 or day 35; Table 2). In a related study, Nottingham (1960) reported low levels of aerobic bacteria in tainted carcasses and concluded that such organisms are most likely not responsible for the condition due to their low population levels. Findings reported by Shank et al. (1962) supported this idea as sour and non-sour knuckles could not be differentiated based on aerobic counts of the muscle tissue, bone marrow, and popliteal lymph nodes. In addition, Shank et al. (1962) aseptically transferred plugs of meat from soured knuckles into knuckles lacking the condition but were not able to induce the souring odor through this process. Based on these findings, Shank et al. (1962) suggested that the condition might not be of bacterial origin.
Mean (n = 10) APC (log CFU/g) and LABC (log CFU/g) for muscle tissue surface samples from knuckles categorized (by plant personnel) as having a slight sour odor, severe sour odor, or no sour odor (control)
| Days of Storage | Bacterial Count Type | Treatment | Mean ± SD | %BDL* |
| 0 | APC | Control | <1.4 ± 0.6a | 20 |
| SLI-SO | 1.6 ± 0.7a | 0 | ||
| SVR-SO | 1.7 ± 0.3a | 0 | ||
| LABC | Control | <1.2 ± 0.5a | 10 | |
| SLI-SO | 1.3 ± 0.4a | 0 | ||
| SVR-SO | <1.3 ± 0.5a | 10 | ||
| 35 | APC | Control | 7.0 ± 0.8a | 0 |
| SLI-SO | 6.9 ± 0.6a | 0 | ||
| SVR-SO | 6.7 ± 1.0a | 0 | ||
| LABC | Control | 7.1 ± 0.7a | 0 | |
| SLI-SO | 7.3 ± 0.9a | 0 | ||
| SVR-SO | 6.7 ± 0.6a | 0 |
Samples were analyzed on the day of collection at the plant (day 0) and after 35 d of refrigerated (0°C ± 2°C) vacuum-packaged storage.
Means with a common superscript letter within each bacterial count type (APC or LABC) and storage day are not (P > 0.05) different. Means with a “less than” symbol (<) indicate that at least one sample within the treatment had a count that was below the analysis detection limit (0.5 log CFU/g).
%BDL indicates the percent of samples, of the 10 samples analyzed, with bacterial counts that were below the analysis detection limit (0.5 log CFU/g).
APC, aerobic plate count; BDL, below detection limit; CFU, colony forming units; LABC, lactic acid bacteria count; SLI-SO, slight sour odor; SVR-SO, severe sour odor.
In the current study, APC and LABC of muscle tissue samples reached approximately 7 log CFU/g after 35 d of vacuum-packaged storage at 0°C ± 2°C (Table 2). The increase in microbial population levels during storage is expected as the low initial numbers of microbial populations recovered on day 0 of storage would be expected to replicate and increase in number through 35 d of refrigerated storage. Recovery of approximately 7 log CFU/g of lactic acid bacteria was an expected result since the product was stored in vacuum packages, and lactic acid bacteria are known to predominate under such packaging conditions (Egan, 1983; James and James, 2002).
Psychrotrophic anaerobic sporeformers in synovial fluid and on femur surface
Low levels of psychrotrophic anaerobic sporeformer populations were recovered from synovial fluid and femur surface sponge samples collected during fabrication (Table 3). Specifically, mean psychrotrophic anaerobic sporeformer counts for synovial fluid samples ranged from <0.8 (SLI-SO and SVR-SO) to <1.0 (control) log CFU/sponge, with no differences (P = 0.88) between the 3 treatment groups (Table 3). Counts of 50% of the control and SLI-SO samples, and 20% of the SVR-SO samples, were below the analysis detection limit (0.4 log CFU/sponge). Similarly, psychrotrophic anaerobic sporeformer counts recovered from sponge samples of the femur surface were low, ranging from <0.8 (SVR-SO) to 1.2 (control) log CFU/sponge (Table 3). Additionally, nondetectable counts (<0.4 log CFU/sponge) were obtained for 40% of the SLI-SO samples and 10% of the SVR-SO samples (Table 3). As seen for the synovial fluid sponge samples, no differences (P > 0.05) were identified between the counts of femur surface swabs taken from control and sour samples (both SLI-SO and SVR-SO). While a few studies have suggested that anaerobic sporeformers—and specifically certain spoilage-causing psychrotrophic Clostridium spp.—could be responsible for the off-odors associated with sour beef rounds (De Lacy et al., 1998), results from the current study could not differentiate (P = 0.88) day 0 sour and non-sour knuckles based on psychrotrophic anaerobic sporeformer contamination within synovial fluid and the femur surface associated with the knuckle. De Lacy et al. (1998) were able to induce the souring condition by inoculating the stifle or hip joints of beef rounds with psychrotrophic Clostridium spp. strains previously isolated from spoiled vacuum-packaged meat products. However, the authors indicated that the high concentration (108 CFU/mL) of Clostridium spp. used for inoculation was not representative of levels of Clostridium spp. populations identified in previous studies (Haines and Scott, 1940; Nottingham, 1960; De Lacy et al., 1998). Furthermore, due to the use of a high inoculation level, the authors were not able to verify whether the off-odors detected were due to growth of the inoculated clostridia or due to presence of existing enzymes within the inoculum suspension (De Lacy et al., 1998). Psychrotrophic Clostridium spp. are also reported to be the causative agents of “blown-pack” spoilage of refrigerated (−1.5°C to 4°C) vacuum-packaged meat products (Adam et al., 2010; Húngaro et al., 2016). This type of spoilage occurs within 2 to 4 wk of processing and is characterized by a putrid odor and severe distension of the packaging due to gas production by the psychrotrophic clostridia (Adam et al., 2010; Húngaro et al., 2016).
Mean (n = 10) psychrotrophic anaerobic sporeformer counts (log CFU/sponge) for synovial fluid sponge samples and femur surface sponge samples collected from knuckles categorized (by plant personnel) as having a slight sour odor, severe sour odor, or no sour odor (control)
| Sample Type | Treatment | Mean ± SD | %BDL* |
| Synovial fluid | Control | <1.0 ± 0.9a | 50 |
| SLI-SO | <0.8 ± 0.6a | 50 | |
| SVR-SO | <0.8 ± 0.4a | 20 | |
| Femur surface | Control | 1.2 ± 0.7a | 0 |
| SLI-SO | <1.2 ± 0.8a | 40 | |
| SVR-SO | <0.8 ± 0.5a | 10 |
Means with a common superscript letter within each sample type analyzed (synovial fluid or femur surface) are not (P > 0.05) different. Means with a “less than” symbol (<) indicate that at least one sample within the treatment had a count that was below the analysis detection limit (0.4 log CFU/sponge).
%BDL indicates the percent of samples, of the 10 samples analyzed, with bacterial counts that were below the analysis detection limit (0.4 log CFU/sponge).
BDL, below detection limit; CFU, colony forming units; SLI-SO, slight sour odor; SVR-SO, severe sour odor.
Overall, the sensory analysis differentiated between sour and non-sour beef knuckles on day 0 and between all 3 categories after 35 d of refrigerated vacuum-packaged storage. However, the culturable microorganisms tested for (i.e., APC, LABC, and psychrotrophic anaerobic sporeformers) were not different among the treatments. This could suggest the possibility that the causative microorganism(s), if the sour condition is in fact of a microbial nature, could be nonculturable. It is estimated that less than 1% of the prokaryotes in most environments can be cultivated in isolation (Schloss and Handelsman, 2005), probably due to the inability to recreate the complex set of environmental conditions required to grow many bacterial species. Therefore, it is possible that the microorganism(s) responsible for producing the sourness could not be detected through a culture-based approach. Culture-independent DNA sequencing techniques could provide a powerful way to rapidly and inexpensively characterize the nonculturable microorganism(s). Specifically, 16S ribosomal RNA (rRNA) sequencing analysis could be utilized to determine whether microbial communities of control (non-sour) and sour knuckles differ (Clarridge, 2004). 16S rRNA sequencing is ideal when working with host-associated samples with high amounts of host DNA (e.g., any meat samples) because a bacterial-specific, taxonomically informative gene is amplified using the polymerase chain reaction and sequenced (Baker et al., 2003; Clarridge, 2004). Unlike shotgun metagenomics, in which all DNA in a sample is sequenced, 16S rRNA sequencing limits DNA sequencing specifically to microbial DNA (Clarridge, 2004; Quince et al., 2017).
The challenges associated with chilling of heavy carcasses could also be a contributing factor for the incidence of sour knuckles. Previous research has demonstrated that the temperature decline in heavier carcasses is slower compared to lighter carcasses (Klauer et al., 2018; Djimsa, 2019). Such issues could be aggravated in deeper muscles leading to a rapid pH decline that can cause the denaturation of proteins (Kim et al., 2014). Previous research evaluating large muscles from the hindquarter of beef, such as the semimembranosus muscle, has demonstrated that the inside region of semimembranosus (the portion closer to the femur) had a higher temperature, lower pH, and a greater abundance of glycolytic enzymes compared to the outside region (Sammel et al., 2002; Nair et al., 2016). With the sour knuckle condition being primarily identified in the internal portion along the femur, it is possible that proper heat dissipation is not occurring. Therefore, the relationship between carcass weight and incidence of sour knuckles needs further investigation.
Conclusions
Sensory panelists were able to identify odor differences between control and sour (SLI-SO and SVR-SO) beef knuckles on the day of sample collection. Furthermore, following 35 d of refrigerated vacuum-packaged storage, the panelists identified odor differences between all 3 treatment groups. However, the microbiological evaluations performed indicated no differences among the treatment groups with regards to APC and LABC of muscle tissue originally in contact with the femur surface, and psychrotrophic anaerobic sporeformer counts of femur surface and synovial fluid sponge samples. It is possible that the sour condition could be due to the presence of microorganisms not tested for in this study, or due to nonculturable microorganisms (organisms that cannot be cultured with traditional methods). The best approach to investigate presence of nonculturable microorganisms would be to conduct a 16S rRNA analysis. There was no statistical difference in the volatiles analyzed using GC-MS, probably due to the gradient of sourness observed in the sour knuckles. The identification of sour knuckles during beef fabrication still remains a challenge, and results from this study could be used as the basis for further research to identify the cause of this quality defect and to develop rapid detection techniques.
Acknowledgements
This research project was partly supported by JBS USA.
Literature Cited
Adam, K. H., S. H. Flint, and G. Brightwell. 2010. Psychrophilic and psychrotrophic clostridia: Sporulation and germination processes and their role in the spoilage of chilled, vacuum-packaged beef, lamb and venison. Int. J. Food Sci. Tech. 45:1539–1544. doi: https://doi.org/10.1111/j.1365-2621.2010.02320.x.
Adhikari, K., L. M. Dooley, E. Chambers, and N. Bhumiratana. 2010. Sensory characteristics of commercial-lactose free milks manufactured in the United States. Lebensm. Wiss. Technol. 43:113–118. doi: https://doi.org/10.1016/j.lwt.2009.06.017.
Baker, G. C., J. J. Smith, and D. A. Cowan. 2003. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Meth. 55:541–555. doi: https://doi.org/10.1016/j.mimet.2003.08.009.
Benjamini, Y., and Y. Hochberg. 1995. Controlling the False Discovery Rate—A practical and powerful approach to multiple testing. J. Roy. Stat. Soc. B. Met. 57:289–300.
Blixt, Y., and E. Borch. 2002. Comparison of shelf life of vacuum-packed pork and beef. Meat Sci. 60:371–378. doi: https://doi.org/10.1016/S0309-1740(01)00145-0.
Boykin, C. A., L. C. Eastwood, M. K. Harris, D. S. Hale, C. R. Kerth, D. B. Griffin, A. N. Arnold, J. D. Hasty, K. E. Belk, D. R. Woerner, R. J. Delmore, J. N. Martin, D. L. VanOverbeke, G. G. Mafi, M. M. Pfeiffer, T. E. Lawrence, T. J. McEvers, T. B. Schmidt, R. J. Maddock, D. D. Johnson, C. C. Carr, J. M. Scheffler, T. D. Pringle, A. M. Stelzleni, J. Gottlieb, and J. W. Savell. 2017. National Beef Quality Audit–2016: In-plant survey of carcass characteristics related to quality, quantity, and value of fed steers and heifers. J. Anim. Sci. 95:2993–3002. doi: https://doi.org/10.2527/jas2017.1543.
Broda, D. M., K. M. De Lacy, and R. G. Bell. 1998. Efficacy of heat and ethanol spore treatments for the isolation of psychrotrophic Clostridium spp. associated with the spoilage of chilled vacuum-packed meats. Int. J. Food. Microbiol. 39:61–68. doi: https://doi.org/10.1016/S01681605(97)00119-0.
Broeckling, C. D., F. A. Afsar, S. Neumann, A. Ben-Hur, and A. Prenni. 2014. RAMClust: A novel feature method enables spectral-matching-based annotation for metabolomics data. Anal. Chem. 14:6812–6917. doi: https://doi.org/10.1021/ac501530d.
Clarridge, J. E. 2004. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin. Microbiol. Rev. 17:840–862. doi: https://doi.org/10.1128/CMR.17.4.840-862.2004.
Corry, J. E. L. 2007. Spoilage organisms of red meat and poultry. In: G. C. Mead, editor, Microbiological analysis of red meat, poultry and eggs. Woodhead Publishing Limited, Cambridge, England. p. 101–122. doi: https://doi.org/10.1533/9781845692513.101.
Cosnett, L. S., D. J. Hogan, N. H. Law, and B. B. Marsh. 1956. Bone-taint in beef. J. Sci. Food Agr. 7:546–551. doi: https://doi.org/10.1002/jsfa.2740070806.
De Lacy, K. M., D. M. Broda, and R. G. Bell. 1998. In vitro assessment of psychrotrophic Clostridium spp. as possible causative agents of bone-taint in beef. Food Microbiol. 15:583–589. doi: https://doi.org/10.1006/fmic.1998.0197.
Djimsa, B. A. 2019. Understanding the impact of carcass size, chilling rate, and electrical stimulation on beef quality. Ph.D. diss. Colorado State University, Fort Collins, CO. (https://mountainscholar.org/handle/10217/195264).
Egan, A. F. 1983. Lactic acid bacteria of meat and meat products. A. Van. Leeuw. J. Microb. 49:327–336. doi: https://doi.org/10.1007/BF00399507.
Gill, C. O. 1979. A review: intrinsic bacteria in meat. J. Appl. Bacteriol. 47:367–378. doi: https://doi.org/10.1111/j.1365-2672.1979.tb01196.x.
Guelker, M. R., A. N. Haneklaus, J. C. Brooks, C. C. Carr, R. J. Delmore, D. B. Griffin, D. S. Hale, K. B. Harris, G. G. Mafi, D. D. Johnson, C. L. Lorenzen, R. J. Maddock, J. N. Martin, R. K. Miller, C. R. Raines, D. L. VanOverbeke, L. L. Vedral, B. E. Wasser, and J. W. Savell. 2013. National Beef Tenderness Survey–2010: Warner-Bratzler shear force values and sensory panel ratings for beef steaks from United States retail and food service establishments. J. Anim. Sci. 91:1005–1014. doi: https://doi.org/10.2527/jas.2012-5785.
Haines, R. B. 1941. The isolation of anaerobes from tainted meats. Chem. Ind.-London. 60:413–416.
Haines, R. B., and W. J. Scott. 1940. An anaerobic organism associated with “bone taint” in beef. J. Hyg. (Lond). 40:154–161. doi: https://doi.org/10.1017/S002217240002773X.
Húngaro, H. M., M. Y. R. Caturla, C. N. Horita, M. M. Furtado, and A. S. Sant’Ana. 2016. Blown pack spoilage in vacuum-packaged meat: A review on clostridia as causative agents, sources, detection methods, contributing factors and mitigation strategies. Trends Food Sci. Tech. 52:123–138. doi: https://doi.org/10.1016/j.tifs.2016.04.010.
Ingram, M. 1952. Internal bacterial taints (“bone taint” or “souring”) of cured pork legs. J. Hyg. (Lond). 50:165–181. doi: https://doi.org/10.1017/S0022172400019525.
James, S. J., and C. James. 2002. Microbiology of refrigerated meat. In: S. J. James, and C. James, editors, Meat refrigeration. Woodhead Publishing Series in Food Science, Technology and Nutrition. Woodhead Publishing Limited, Cambridge, England. p. 3–19. doi: https://doi.org/10.1016/9781855736535.1.3.
Kim, Y. H. B., R. D. Warner, and K. Rosenvold. 2014. Influence of high pre-rigor temperature and fast pH fall on muscle proteins and meat quality: A review. Anim. Prod. Sci. 54:375–395. doi: https://doi.org/10.1071/AN13329.
Klauer, B. L. 2018. Mapping temperature decline in beef cattle during conventional chilling. M.S. thesis. Colorado State University, Fort Collins, CO. (https://mountainscholar.org/handle/10217/195237).
Lepovetsky, B. C., H. H. Weiser, and F. E. Deatherage. 1953. A microbiological study of lymph nodes, bone marrow and muscle tissue obtained from slaughtered cattle. J. Appl. Microbiol. 1:57–59.
McKenna, D. R., D. L. Roebert, P. K. Bates, T. B. Schmidt, D. S. Hale, D. B. Griffin, J. W. Savell, J. C. Brooks, J. B. Morgan, T. H. Montgomery, K. E. Belk, and G. C. Smith. 2002. National Beef Quality Audit-2000: Survey of targeted cattle and carcass characteristics related to quality, quantity, and value of fed steers and heifers. J. Anim. Sci. 80:1212–1222. doi: https://doi.org/10.2527/2002.8051212x.
Nair, M. N., S. P. Suman, M. K. Chatli, S. Li, P. Joseph, C. M. Beach, and G. Rentfrow. 2016. Proteome basis for intramuscular variation in color stability of beef semimembranosus. Meat. Sci. 113:9–16. doi: https://doi.org/10.1016/j.meatsci.2015.11.003.
Nottingham, P. M. 1960. Bone-taint in beef—II. Bacteria in ischiatic lymph nodes. J. Sci. Food Agr. 11:436–441. doi: https://doi.org/10.1002/jsfa.2740110803.
Pérez, R. A., M. D. Rojo, González, G., and C. De Lorenzo. 2008. Solid-phase microextraction for the determination of volatile compounds in the spoilage of raw ground beef. J. AOAC Int. 91:1409–1415.
Quince, C., A. W. Walker, J. T. Simpson, N. J. Loman, and N. Segata. 2017. Shotgun metagenomics, from sampling to analysis. Nat. Biotechnol. 35:833–844. doi: https://doi.org/10.1038/nbt.3935.
Sammel, L. M., M. C. Hunt, D. H. Kropf, K. A. Hachmeister, C. L. Kastner, and D. E. Johnson. 2002. Influence of chemical characteristics of beef inside and outside semimembranosus on color traits. J. Food Sci. 67:1323–1330. doi: https://doi.org/10.1111/j.1365-2621.2002.tb10282.x.
Savell, J. W., S. L. Mueller, and B. E. Baird. 2005. The chilling of carcasses. Meat Sci. 70:449–459. doi: https://doi.org/10.1016/j.meatsci.2004.06.027.
Schloss, P. D., and J. Handelsman. 2005. Metagenomics for studying unculturable microorganisms: Cutting the Gordian knot. Genome Biol. 2005;6(8):299 doi: https://doi.org/10.1186/gb-2005-6-8-229.
Scott, V. N., J. E. Anderson, and G. Wang. 2001. Mesophilic anaerobic sporeformers. In: F. P. Downes, and I. Keith, editors, Compendium of methods for the microbiological examination of food. 4th ed. American Public Health Association, Washington, DC. p. 229–235.
Shank, J. L., J. H. Silliker, and P. A. Goeser. 1962. The development of a nonmicrobial off-condition in fresh meat. Appl. Environ. Microb. 10:240–246.
Smith, C. A., E. J. Want, G. O’Maille, R. Abagyan, and G. Siuzdak. 2006. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 78:779–787. doi: https://doi.org/10.1021/ac051437y.